Sự gia tăng dân số đã tạo nên áp lực lớn đối với nền nông nghiệp trong việc đảm bảo an ninh lương thực. Trong khi đó, với tốc độ biến đổi khí hậu như ngày này, việc mất mùa đang diễn ra liên tục trên thế giới. Một trong những nguyên nhân đó chính là sự kém thích nghi kịp thời của thực vật đối với quá trình biến đổi khí hậu.
Trong các thập kỷ gần đây, stress phi sinh học trở thành mối quan tâm chính trong sản xuất nông nghiệp. Sự tăng trưởng và phát triển của thực vật trong các điều kiện stress có thể ảnh hưởng đến vụ mùa. Trong các điều kiện stress, thực vật vẫn có những cơ chế để thích nghi đối với sự thay đổi cường độ khác nhau của stress, tuy nhiên những cơ chế này vẫn chưa đủ để giúp thực vật chống chịu. Hiện nay, Abscisic Acid (ABA) được sử dụng để giúp thực vật thích nghi tốt hơn trước những thay đổi của môi trường và khí hậu hiện nay. Vậy ABA đã giúp thực vật thích nghi như thế nào?
* Stress phi sinh học là các điều kiện khắc nghiệt như: nhiệt độ, nước, nồng độ muối…
VAI TRÒ CỦA ABA ĐỐI VỚI THỰC VẬT
Hormone thực vật là chìa khóa trong điều hòa quá trình tăng trưởng và phát triển cũng như đáp ứng với các điều kiện môi trường khác nhau, trong đó có Abscisic Acid (ABA). Khi điều kiện môi trường trở nên khắc nghiệt, thực vật tăng cường tổng hợp ABA. ABA tương tác với các thụ thể để kích hoạt các con đường tín hiệu giúp tế bào đáp ứng với stress, chính vì thế, ABA còn được gọi là hormone stress (1).
Hạt là một cơ quan quan trọng trong sự phát triển của các thực vật bậc cao, quá trình thay đổi từ giai đoạn ngủ đến quá trình nảy mầm là giai đoạn quan trọng trong chu trình sống và được xem như là một đặc điểm sinh thái và dấu vết thích nghi có ở thực vật. Hai hormone đóng vai trò trung tâm trong kiểm soát quá trình ngủ đông và nảy mầm là ABA và gibberellins (GAs). Cả hai hormone này đều kiểm soát sự cân bằng giữa giai đoạn ngủ và giai đoạn nảy mầm. ABA đóng vai trò trong việc cảm ứng và duy trì trạng thái ngủ của hạt. Trong suốt giai đoạn khô, việc chuyển hóa ABA cần phải được điều hòa.
Đối với hệ thống rễ, các stress phi sinh học diễn ra khi sự khan hiếm nước xuất hiện, hoặc môi trường nước không đồng đều (lúc có lúc không). Trong các trường hợp này, hàm lượng ABA thay đổi để đáp ứng với stress nước. Kết quả là sự thay đổi môi trường ở rễ sẽ ảnh hưởng đến sự đáp ứng ABA tại vùng rễ toàn bộ cây (2).
Khí khổng là những lỗ khí nhỏ ở bề mặt lá được hình thành bởi các tế bào bảo vệ có chức năng kiểm soát sự trao đổi khí. Ánh sáng có khả năng kích thích sự mở khí khổng. Trong khi đó, ABA và hàm lượng CO2 kích thích sự đóng khí khổng. Trong quá trình đóng khí khổng, việc giảm trao đổi khí có thể dẫn đến giảm năng suất quang hợp và việc mất hơi nước từ lá có thể giảm. Trong điều kiện khô hạn, sự thay đổi ABA đảm bảo sự trao đổi ion ở các tế bào bảo vệ, kích thích việc đóng khí khổng và ngăn khí khổng mở và làm giảm tình trạng mất nước (3).
TỔNG HỢP, XÚC TÁC VÀ VẬN CHUYỂN ABA
Tổng Hợp ABA
ABA được bắt đầu tổng hợp ở các bào quan nhỏ và lưu trữ trong tế bào chất. ABA được tổng hợp ở trong tế bào thực vật bậc cao thông qua con đường phụ thuộc vào mevalonic acid hay còn gọi là con đường gián tiếp. Trong con đường này, ABA được tổng hợp bằng sự cắt bỏ C40 của tiền carotenoid, bằng cách chuyển đổi hai bước xanthoxin trung gian sang ABA thông qua aldehyde ABA và được oxy hóa thành ABA (Hình 1).

Bước đầu tiên của con đường tổng hợp ABA là chuyển đổi tất cả các trans-violaxanthin thành zeaxanthin và được xúc tác bởi zaxanthin epoxidase (ZEP) trong plastid. Trong phản ứng này, antheraxanthin là chất trung gian được hình thành. Sau đó, tất cả trans-violaxanthin chuyển thành 9-cis-violaxanthin hoặc 9-cis-neoxanthin. Tiếp theo, enzyme 9-cis-epoxy carotenoid dioxygenase (NCED) xúc tác phân hủy oxy hoá 9-cis-violaxanthin và 9-cis-neoxanthin sinh ra sản phẩm trung gian C15 gọi là xanthoxin và chất chuyển hóa C25. Cuối cùng, xanthoxin được xuất ra cytosol, nơi xanthoxin được chuyển thành ABA.
Xúc Tác ABA
Khi tín hiệu stress giảm, ABA được chuyển hóa thành các sản phẩm không hoạt động (1). Quá trình này được thực hiện bởi hai con đường: hydroxyl hóa và liên hợp (4). Trong hydroxyl hóa, ABA được hydroxyl hóa qua quá trình oxy hóa của ba nhóm methyl (C-7’, C-8’, và C-9’) trong cấu trúc vòng.
ABA và hoạt động xúc tác hydroxyl của ABA có thể được liên hợp với glucose. ABA glucosyl este (ABA-GE) được tổng hợp bằng glycosyltransferase và được lưu trữ trong không bào. Trong điều kiện stress phi sinh học, ABA glucosyl ester có thể được chuyển đổi thành ABA bằng thủy phân xúc tác enzyme. Enzyme glycosidase xúc tác sự thủy phân của ABA-GE để giải phóng.
Vận Chuyển ABA
Sự vận chuyển ABA giữa các tế bào, các mô và các cơ quan cũng đóng vai trò quan trọng trong phản ứng sinh lý toàn bộ thực vật trong điều kiện stress. ABA, là một axit yếu, có thể khuếch tán thụ động qua màng sinh học khi ABA được proton hóa (5). ABA cũng có thể được vận chuyển qua màng bởi chất vận chuyển.
TÍN HIỆU ABA
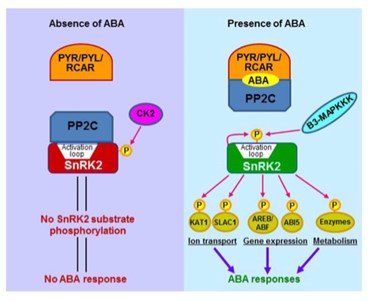
Abscisic Acid là tín hiệu nội bào quan trọng trong thực vật và có vai trò rất quan trọng trong các đáp ứng stress khác nhau ở thực vật. Do đó, sự hiểu biết cơ chế tín hiệu của ABA rất quan trọng để cải thiện năng suất của cây trồng trong điều kiện stress và rất cần thiết khi khí hậu đang ngày càng trở nên nóng hơn. Có ba thành phần chính tham gia tín hiệu ABA: Kháng pyralin / kháng pyrabactin (PYL) / thành phần quy định của thụ thể ABA (RCAR), protein phosphatase 2C (PP2C: hoạt động như các điều hòa âm tính) và (Sucrose không lên men) SNF1 protein kinase 2 (SnRK2: Hoạt động như một chất điều hòa dương tính). Với sự có mặt của ABA, phức hợp PYR / PYL / RCAR-PP2C được hình thành và dẫn đến sự ức chế hoạt động của PP2C, cho phép kích hoạt SnRK2. Sự kích hoạt SnRK2 làm cho các protein ở hạ nguồn được phospho hóa và hoạt động như các yếu tố phiên mã, và do đó tạo điều kiện sao chép các gen cần cho việc đáp ứng ABA (Hình 2).
SỰ ĐÁP ỨNG TÍNH HIỆU ABA Ở MỨC BIỂU HIỆN GENE
Kiểm Soát Phiên Mã Điều Hòa Bởi ABA
Các yếu tố phiên mã đóng vai trò trong điều hòa, cân bằng và điều phối các tín hiệu nội tiết, phát triển trong các hệ thống thực vật (6). Một nhân tố phiên mã có thể điều chỉnh sự biểu hiện của nhiều gen mục tiêu thông qua liên kết với một nhân tố phiên mã khác bởi các yếu tố cis trong promoter của các gen đích tương ứng. Bộ gen thực vật chỉ có khoản 7% trình tự mã hóa cho các yếu tố phiên mã (7).
Hạn hán và stress muối hoạt động như một tín hiệu stress và dẫn đến sự tích lũy ABA (1). Tín hiệu ABA làm thay đổi đáng kể sự biểu hiện gen, dẫn đến sự thay đổi trong quá trình phiên mã, dịch mã, và ổn định. Biểu hiện gen phụ thuộc vào ABA cần có sự liên kết của các yếu tố phiên mã với các yếu tố điều kiện cis (cis-acting elements) trong các promoter gen mục tiêu. Các kinase kích hoạt do ABA chịu trách nhiệm phosphoryl hoá các yếu tố phụ thuộc ABA (8).
Kiểm Soát Sau Dịch Mã Trong Điều Hòa ABA
Các protein liên kết RNA có vai trò quan trọng trong kiểm soát sự biểu hiện gene sau quá trình phiên mã, bao gồm: cắt nối mRNA (spicing), sự ổn định của mRNA, sự định vị của mRNA và dịch mã (9). Các protein liên kết RNA đóng vai trò quan trọng trong việc đáp ứng với ABA và thường có nhiều domain bám có khả năng cảm ứng sự nhận diện motif, như K homology domain, zinc finger, cold-shock domain, aspartate- glutamate-alanine-aspartate (DEAD) box, and double-stranded RNA-binding domain (10).
Bên cạnh quá trình biến đổi sau dịch mã, việc kiểm soát bằng ubiquitin đóng vai trò quan trọng trong tín hiệu ABA. Quá trình Ubiquitin hóa được thực hiện bởi các emzyme như: ubiquitin-activating enzymes (E1s), ubiquitin conjugating enzymes (E2s), và ubiquitin ligases (E3s). Quá trình ubiquitin hóa có thể ảnh hưởng đến protein theo nhiều cách khác nhau: gắn đuôi lên protein để phân hủy protein nhờ proteasome, thay đổi vị trí của protein trong tế bào, thúc đẩy hoặc ngăn cản sự tương tác giữa các protein. Những protein quan trọng trong tín hiệu của ABA được gắn ubiquitin cần cho sự phân giải proteasome. Quá trình này có thể được cảm ứng khi ABA ở nồng độ thấp. Sự phân giải các receptor của ABA do proteasome cho phép sự giải phóng PP2C, điều hòa âm tính SnRK2. Hay nói cách khác, khi ABA ở hàm lượng cao thì quá trình trên bị ức chế (11).
Biến Đổi Ngoại Gen (Epigenetics) Trong Điều Hòa Để Đáp Ứng ABA
Biến đổi ngoại gen trong chức năng của gen không phải do sự thay đổi trình tự nucleotide DNA, mà là do sự biến đổi hóa học của DNA và các protein liên quan của nó. Các biến đổi ngoại gen bao gồm sự methyl hóa DNA, sửa đổi histone và sản xuất microRNA. Vì các yếu tố stress môi trường có thể gây ra sự thay đổi ngoại gen của bộ gen, dẫn đến cơ chế quan trọng trung gian giữa sự tương tác giữa gen và môi trường (12).
Trong số các sửa đổi histone, acetyl hóa histone đã được xem là quan trọng trong việc điều chỉnh các gen đáp ứng ABA và stress. Deacetylation histone đóng vai trò thiết yếu trong việc kích hoạt các gen đáp ứng ABA để thích nghi hạn hán. Ngược lại, đột biến hoặc ức chế gen histamine deacetylase histone làm giảm biểu hiện của ABA và các gen đáp ứng với stress phi sinh học trong đó có stress muối.
Lời kết
Quá trình biến đổi khí hậu đang diễn tiến ngày càng phức tạp cùng sự gia tăng dân số đang trở thành áp lực to lớn đối với nền nông nghiệp thế giới. Ngày càng nhiều các nghiên cứu đang được tiến nhằm cải thiện năng suất và gia tăng sức chống chịu của cây trồng. Acid Abscisic là một trong những đối tượng nghiên cứu quan trọng để cải thiện các giống lương thực hiện nay. Bên cạnh đó, còn nhiều hormone thực vật khác cũng đang được nghiên cứu. Vì thế, chúng ta có thể hy vọng rằng trong tương lai gần, an ninh lương thực thế giới vẫn có thể được duy trì. Tuy nhiên, song song với việc nghiên cứu để tạo ra nhiều giống cây trồng ưu việt thì việc nâng cao ý thức con người nhằm hạn chế tối đa tốc độ biến đổi khí hậu cũng là một khía cạnh quan trọng.
Tài liệu tham khảo:
1. Ng, L. M., Melcher, K., Teh, B. T., and Xu, H. E., “Abscisic acid perception and signaling: structural mechanisms and applications.,” cta Pharmacol., vol. 35, p. 567–584, 2014.
2. Dodd, I. C., Egea, G., and Davies, W. J., “Abscisic acid signaling when soil moisture is heterogeneous: decreased photoperiod sap flow from drying roots limits abscisic acid export to the shoots,” Plant Cell Environ, pp. 31, 1263–1274, 2008.
3. Aswath, C. R., Kim, S. H., Mo, S. Y., and Kim, D. H., “Transgenic plants of creeping bent grass harboring the stress-inducible gene, 9-cis-epoxycarotenoid dioxygenase, are highly tolerant to drought and NaCl Stress,” Plant Growth Regul, pp. 47, 129–139, 2005.
4. Nambara, E., and Marion-Poll, “Abscisic acid biosynthesis and catabolism,” Annu. Rev. Plant Biol, pp. 56, 165–185, 2005.
5. Wilkinson, S., and Davies, W. J., “Drought, ozone, ABA and ethylene: new insights from cell to plant to community,” Plant Cell Environ., pp. 33, 510–525, 2010.
6. Jaradat, M. R., Feurtado, J. A., Huang, D., Lu, Y., and Cutler, A. J., “Multiple roles of the transcription factor AtMYBR1/AtMYB44 in ABA signaling, stress responses, and leaf senescence,” BMC Plant Biol, 2013.
7. Udvardi, M. K., Kakar, K., Wandrey, M., Montanari, O., Murray, J., Andriankaja, A., et al, “Legume transcription factors: global regulators of plant development and response to the environment,” Plant Physiol, pp. 144, 538–549, 2007.
8. Johnson, R. R., Wagner, R. L., Verhey, S. D., and Walker-Simmons, M. K. , “The abscisic acid-responsive kinase PKABA1 interacts with a seed-specific abscisic acid response element-binding factor, TaABF, and phosphorylates TaABF peptide sequences.,” Plant Physiol, p. 837–846, 2002.
9. Glisovic, T., Bachorik, J. L., Yong, J., and Dreyfuss, G. , “NA-binding proteins and post-transcriptional gene regulation. FEBS Lett. 582, 1977–1986. doi: 10.1016/j.febslet.2008.03.004,” FEBS Lett, p. 1977–1986, 2008.
10. Ambrosone, A., Costa, A., Leone, A., and Grillo, S., “Beyond transcription: RNA-binding proteins as emerging regulators of plant response to environmental constraints,” Plant Sci, p. 12–18, 2012.
11. Irigoyen, M. L., Iniesto, E., Rodriguez, L., Puga, M. I., Yanagawa, Y., Pick, E., “Targeted degradation of abscisic acid receptors is mediated by the ubiquitin ligase substrate adaptor DDA1 in Arabidopsis,” Plant Cell, p. 712–728, 2014.
12. G. J. King, “Crop epigenetics and the molecular hardware of genotype × environment interactions,” Front Plant Sci, p. 968, 2015.
13. Saroj K.Sah, Kambham R.Reddy, Jiaxu Li, “Abscisic Acid and Abiotic Stress Tolerance in Crop Plants,” Plant Sci, 04 May 2016 .
Tác giả: Nguyễn Thị Tuyết
Trường Đại Học Khoa Học Tự Nhiên, TP Hồ Chí Minh
Theo VJS



