tung.biotech
Senior Member
[FONT="]Protein bất hoạt ribosome ở thực vật[/FONT]
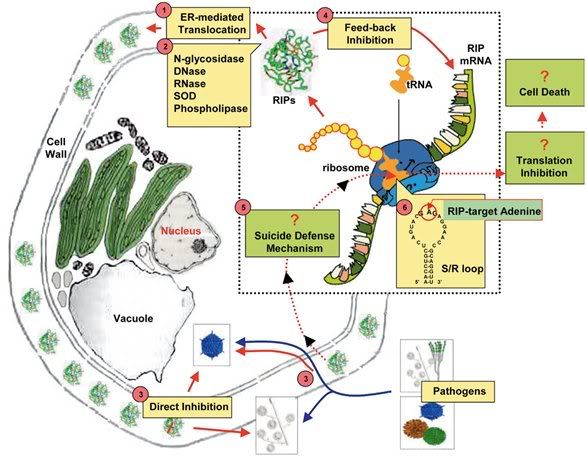
[FONT="]- Ribosome-inactivating protein (RIPs) là một nhóm các chất độc tế bào (cytotoxic) N-glycosidases mà cụ thể là tách mối liên kết N-C glycosidic của nucleotide. RIPs đã được phân loại vào ba loại: type I gồm có một chuỗi polypeptide đơn, trong khi type II là một heterodimer bao gồm một chuỗi A chức năng tương đương với type I, chuỗi B gắn liên kết đường . Type III là một chuỗi đơn chứa vùng carboxyl-terminal mở rộng chưa biết chức năng. Mặc dù RIP đã được xác định lần đầu tiên cách đây hơn 100 năm về trước, các chức năng sinh học vẫn còn đang mở để nghiên cứu. Ngoài ra, những hiểu biết về hoạt động loại purin (depurinating) ribosome in vitro tại vòng sarcin/ricin (S/R) chưa trả lời rõ ràng cho câu hỏi tại sao thực vật tổng hợp và tích lũy RIP. Báo cáo này đề cập những tiến bộ gần đây trên RIPs và đề xuất một mô hình làm việc trong một cố gắng để tổng kết và hợp lý hóa chức năng sinh học trong cây.[/FONT]
- [FONT="]Một lý do quan trọng của chúng ta về những không hiểu biết về RIPs là thiếu thông tin về nhóm này của enzymes từ một mô hình hệ thống cây trồng nghiên cứu tốt, chẳng hạn như Arabidopsis thaliana hay Nicotiana tabacum. Nó đã được suy nghĩ RIPs không được tìm thấy phổ biến giữa các cây trồng, bằng chứng không xác định RIPs trong các mô hình hệ thống cây trồng. Gần đây, tuy nhiên,[/FONT][FONT="] , a 26-kDa RIP-like protein termed TRIP[/FONT][FONT="] , có N-glycosidase hoạt động, đã được tách chiết và mô tả từ N. tabacum (Sharma et al. 2004). Trip đã được tìm thấy và biểu hiện ở mức độ rất thấp trong lá (0.01% tổng nguyên liệu ban đầu), mà có thể là lý do đằng sau một thời gian dài không thành công khi xác định RIP từ thuốc lá. Khám phá này mở ra khả năng loài thực vật khác cũng có một hay nhiều N-glycosidases. [/FONT]
- [FONT="]Gần đây, ngày càng tăng tranh cãi là RIPs có hay không hoạt tính enzyme khác ngoài hoạt tính N-glycosidase. Hầu hết các khám phá gần đây về hoạt tính enzyme của RIPs là hoạt tính RNase và DNase. Ngoài ra, một số RIPs có hoạt tính superoxide dismutase (SOD, Li et al. 1997) và phospholipase (Helmy et al. 1999). Ban đầu những hoạt enzymatic mới đã không được để ý đến, và nghĩ rằng do không tinh khiết trong việc chuẩn bị của RIPs (Day et al. 1998; Valbonesi et al. 1999). Tuy nhiên, hoạt tính DNase và/hoặc đã được kiểm tra lại và xác nhận bằng cách sử dụng pokeweed antiviral protein (PAP) tinh khiết cao và một chuỗi A cinnamomin tái tổ hợp (từ Cinnamomum camphora; Wang và Tumer 1999; He và Liu 2004). Calcaelin (từ Calvatia caelata), tương tự RIPs thực vật, cũng đã được biểu lộ hoạt tính RNase (Ng et al. 2003). Hoạt tính SOD được báo cáo cho camphorin (từ C. camphora) chưa được độc lập xác nhận, nhưng, điều thú vị, TRIP cho thấy hoạt tính SOD, thêm vào các hoạt tính N-glycosidase (Sharma et al. 2004). Ngược lại, Fe-SOD cô lập từ Escherichia coli đã được cũng được tìm thấy có hoạt tính N-glycosidase lên ribosomes nấm men cùng với các hoạt tính superoxide-scavenging (Sharma et al. 2004), chứng tỏ rõ ràng rằng một vài RIPs cho thấy hoạt tính SOD không thể là kết quả của nhiễm bẩn. Có vẻ là cần phải xem xét tất cả các dữ liệu về các protein để hiểu đầy đủ các chức năng sinh học của họ. [/FONT]
- [FONT="]RIP được biết có vai trò quan trọng sự phòng thủ của thực vật kể từ khi dịch chiết thô của lá cây pokeweed (cây thương lục Mỹ) lần đầu tiên cho thấy hoạt tính ức chế chống lại sự sâm nhiễm virus vào cây. Kể từ đó, một vài phương pháp công nghệ sinh học có cố gắn để tạo ra cây chuyển gen để khai thác hoạt tính chống vi khuẩn của RIP. Biểu hiện một vài RIP (PAP, trichosanthin từ cây trichosanthes kirilowii và protein kháng vivus từ cây Phytolacca insularis) trong cây chuyển gen dẫn đến thành công trong việc kháng cự chống lại nhiều loại virus khác nhau và/hoặc mần bệnh nấm (Lodge et al. 1993; Lam et al. 1996; Moon et al. 1997). Việc thừa nhận rộng rãi cơ chế kháng vi sinh vật hoạt động nhận dạng ribosome vật chủ với mục tiêu của hoạt tính RIP. Mầm bệnh lây nhiễm được nghĩ là thay đổi cấu trúc bên trong của tế bào vật chủ, cho phép RIP sử dụng ribosome và chỉ huy dừng việc tổng hợp protein và tế bào chết (Ready et al. 1986). Tuy nhiên, không có bằng chứng nào ủng hộ cơ chế tự sát. Một vài thí nghiệm cây chuyển gen biểu hiện RIP có cho thấy rằng cây không tỏ ra triệu chứng tế bào chết tự nhiên trong phản ứng lại sự xâm nhập của vi sinh vật mặt dù chúng kháng lại phạm vi lớn của mầm bệnh.[/FONT]
- [FONT="]Một số nghiên cứu độc lập đề xuất rằng các hoạt động kháng vi sinh vật của RIP được tách ra khỏi hoạt động ức chế ribosome vật chủ (host-ribosome-inactivating – RI). Chứng minh sớm nhất được thực hiện bởi Tumer và đồng nghiệp của cô, họ thí nghiệm đột biến xóa của PAP trong cây thuốc lá chuyển gen (Zoubenko et al. 1997). Mặt dầu PAPc (PAPW327stop) và PAPn (PAPG75D) không thể loại purin tại vòng S/R, chúng vẫn kháng được mầm bệnh thực vật. Gần đây, cơ chế ngăn chặn của RIP chống lại virus suy giảm miễn dịch ở người (human immunodeficiency virus - HIV) tái bản, mọi người đều tin rằng có liên quan đến hoạt động RI, cũng đã được readdress (Wang et al. 2003). Phương pháp đột biến điểm định hướng (Site-directed mutagenesis) sử dụng trichosanthin đã chứng minh được hoạt động RI cổ điển giải thích không đầy đủ hoạt đọng kháng HIV. Trichosanthin đột biến xóa C-terminal hay thêm amino acid giữ lại hầu hết tất cả hoạt động RI, nhưng không có hoạt động kháng HIV. Hơn nửa, ngày càng tăng bằng chứng chứng minh rằng RIP có khả năng nhận ra và hành động ngay lập tức trên virus và acid nucleotid cảm ứng virus, hủy bỏ những lây nhiêm của chúng (Rajamohan et al. 1999; Park et al. 2004). Chúng cũng được thông báo rằng hoạt động RI của RIP không được coi như chất độc tế bào (Park et al. 2002). Những kết quả này (Hình 1) cho biết (i) hoạt động enzyme của RIP chưa giải thích đầy đủ hành động bảo vệ chống lại mầm bệnh và (ii) cơ chế bảo vệ có thể bắt đầu bằng trực tiếp lên mầm bệnh xâm nhiễm mục tiêu hơn là ribosome vật chủ.[/FONT]
- [FONT="]Vai trò sinh học khác có thể của RIPs thu được từ bằng chứng ngày càng tăng rằng chúng có thể nhắm mục tiêu không phải ribosome. Phần còn lại adenine bảo tồn phổ biến A4324 trong vòng S/R trong một thời gian dài được coi là vị trí mục tiêu duy nhất của RIP, nhưng nghiên cứu gần đây phát hiện có thể thay thế khác vòng S/R. ví dụ, một số RIP cho thấy có khả năng giải phóng Adenine từ nhiều vị trí trong rRNA và nucleic acid poly(A), mRNA, tRNA, rRNA, và DNA (Barbieri et al. năm 1992, 1997; Park et al. 2004). Một vài RIP còn biểu lộ hoạt động chống lại poly(ADP-ribosyl)ated poly(ADP-ribose) polymerase (enzyme tự động sửa đổi; Barbieri et al. 2003). Khả năng này ảnh hưởng đến các chất khác cho phép RIP điều hòa chức năng quang trọng của tế bào[/FONT]
- [FONT="]RIP có thể điều chỉnh biểu hiện protein bằng mRNA mục tiêu thay cho ribosome. PAP cho thấy có thể ảnh hưởng trở lại trong biểu hiện chính mRNA của chúng trong nấm men chuyển gen (Parikh et al. 2002). Biểu hiện bên ngoài của PAP dẩn đến ổn định của chính mRNA của nó sau khi mức độ mRNA được điều chỉnh lên bằng cơ chế xảy ra độc lập với loại purine rRNA và dịch mã. Trong hệ thống thực vật, gene ME1 cảm ứng bằng xử lý jasmonic acid, nhưng mức độ phiên mã cao gene ME1 không có kết quả trong dịch mã thành protein in vitro (Vepachedu et al. 2003). Trong nghiên cứu in vitro cho thấy ME1 có thể loại purine chính mRNA của chúng và theo cách đó ức chế dịch mã chính nó. Bởi vậy, có thể rằng RIP có khả năng điều hòa mức độ biểu hiện của chính nó.[/FONT]
- [FONT="]Mặc dù đã được nghiên cứu tỉ mỹ từ lâu dài như là một độc chất hữu ích tiềm năng, vai trò và chức năng của RIP trong thực vật và sự phân bố tự nhiên vẫn còn khó hiểu. việc nhận dạng RIP từ cơ thể mô hình N. tabacum, chung với genomic và phương pháp công nghệ sinh học, vhawngr hạn như mutagenesis và gene silencing, có thể là một cách hiệu quả lấp lỗ hổng kiến thức. trong một vài năm tới, nó có khả năng phát triển nhanh trong lĩnh vực này. Dẫn đầu là làm sáng tỏ chức năng sinh học của RIP và chứng minh vai trò coe bản trong tế bào thực vật và tiềm năng đáng kể của chúng trong một vài ứng dụng chữa bệnh bao gồm chuẩn bị độc chất miễn dịch mục tiêu khối u và u máu ác tính. [/FONT]
[FONT="]- Ribosome-inactivating protein (RIPs) là một nhóm các chất độc tế bào (cytotoxic) N-glycosidases mà cụ thể là tách mối liên kết N-C glycosidic của nucleotide. RIPs đã được phân loại vào ba loại: type I gồm có một chuỗi polypeptide đơn, trong khi type II là một heterodimer bao gồm một chuỗi A chức năng tương đương với type I, chuỗi B gắn liên kết đường . Type III là một chuỗi đơn chứa vùng carboxyl-terminal mở rộng chưa biết chức năng. Mặc dù RIP đã được xác định lần đầu tiên cách đây hơn 100 năm về trước, các chức năng sinh học vẫn còn đang mở để nghiên cứu. Ngoài ra, những hiểu biết về hoạt động loại purin (depurinating) ribosome in vitro tại vòng sarcin/ricin (S/R) chưa trả lời rõ ràng cho câu hỏi tại sao thực vật tổng hợp và tích lũy RIP. Báo cáo này đề cập những tiến bộ gần đây trên RIPs và đề xuất một mô hình làm việc trong một cố gắng để tổng kết và hợp lý hóa chức năng sinh học trong cây.[/FONT]
- [FONT="]Một lý do quan trọng của chúng ta về những không hiểu biết về RIPs là thiếu thông tin về nhóm này của enzymes từ một mô hình hệ thống cây trồng nghiên cứu tốt, chẳng hạn như Arabidopsis thaliana hay Nicotiana tabacum. Nó đã được suy nghĩ RIPs không được tìm thấy phổ biến giữa các cây trồng, bằng chứng không xác định RIPs trong các mô hình hệ thống cây trồng. Gần đây, tuy nhiên,[/FONT][FONT="] , a 26-kDa RIP-like protein termed TRIP[/FONT][FONT="] , có N-glycosidase hoạt động, đã được tách chiết và mô tả từ N. tabacum (Sharma et al. 2004). Trip đã được tìm thấy và biểu hiện ở mức độ rất thấp trong lá (0.01% tổng nguyên liệu ban đầu), mà có thể là lý do đằng sau một thời gian dài không thành công khi xác định RIP từ thuốc lá. Khám phá này mở ra khả năng loài thực vật khác cũng có một hay nhiều N-glycosidases. [/FONT]
- [FONT="]Gần đây, ngày càng tăng tranh cãi là RIPs có hay không hoạt tính enzyme khác ngoài hoạt tính N-glycosidase. Hầu hết các khám phá gần đây về hoạt tính enzyme của RIPs là hoạt tính RNase và DNase. Ngoài ra, một số RIPs có hoạt tính superoxide dismutase (SOD, Li et al. 1997) và phospholipase (Helmy et al. 1999). Ban đầu những hoạt enzymatic mới đã không được để ý đến, và nghĩ rằng do không tinh khiết trong việc chuẩn bị của RIPs (Day et al. 1998; Valbonesi et al. 1999). Tuy nhiên, hoạt tính DNase và/hoặc đã được kiểm tra lại và xác nhận bằng cách sử dụng pokeweed antiviral protein (PAP) tinh khiết cao và một chuỗi A cinnamomin tái tổ hợp (từ Cinnamomum camphora; Wang và Tumer 1999; He và Liu 2004). Calcaelin (từ Calvatia caelata), tương tự RIPs thực vật, cũng đã được biểu lộ hoạt tính RNase (Ng et al. 2003). Hoạt tính SOD được báo cáo cho camphorin (từ C. camphora) chưa được độc lập xác nhận, nhưng, điều thú vị, TRIP cho thấy hoạt tính SOD, thêm vào các hoạt tính N-glycosidase (Sharma et al. 2004). Ngược lại, Fe-SOD cô lập từ Escherichia coli đã được cũng được tìm thấy có hoạt tính N-glycosidase lên ribosomes nấm men cùng với các hoạt tính superoxide-scavenging (Sharma et al. 2004), chứng tỏ rõ ràng rằng một vài RIPs cho thấy hoạt tính SOD không thể là kết quả của nhiễm bẩn. Có vẻ là cần phải xem xét tất cả các dữ liệu về các protein để hiểu đầy đủ các chức năng sinh học của họ. [/FONT]
- [FONT="]RIP được biết có vai trò quan trọng sự phòng thủ của thực vật kể từ khi dịch chiết thô của lá cây pokeweed (cây thương lục Mỹ) lần đầu tiên cho thấy hoạt tính ức chế chống lại sự sâm nhiễm virus vào cây. Kể từ đó, một vài phương pháp công nghệ sinh học có cố gắn để tạo ra cây chuyển gen để khai thác hoạt tính chống vi khuẩn của RIP. Biểu hiện một vài RIP (PAP, trichosanthin từ cây trichosanthes kirilowii và protein kháng vivus từ cây Phytolacca insularis) trong cây chuyển gen dẫn đến thành công trong việc kháng cự chống lại nhiều loại virus khác nhau và/hoặc mần bệnh nấm (Lodge et al. 1993; Lam et al. 1996; Moon et al. 1997). Việc thừa nhận rộng rãi cơ chế kháng vi sinh vật hoạt động nhận dạng ribosome vật chủ với mục tiêu của hoạt tính RIP. Mầm bệnh lây nhiễm được nghĩ là thay đổi cấu trúc bên trong của tế bào vật chủ, cho phép RIP sử dụng ribosome và chỉ huy dừng việc tổng hợp protein và tế bào chết (Ready et al. 1986). Tuy nhiên, không có bằng chứng nào ủng hộ cơ chế tự sát. Một vài thí nghiệm cây chuyển gen biểu hiện RIP có cho thấy rằng cây không tỏ ra triệu chứng tế bào chết tự nhiên trong phản ứng lại sự xâm nhập của vi sinh vật mặt dù chúng kháng lại phạm vi lớn của mầm bệnh.[/FONT]
- [FONT="]Một số nghiên cứu độc lập đề xuất rằng các hoạt động kháng vi sinh vật của RIP được tách ra khỏi hoạt động ức chế ribosome vật chủ (host-ribosome-inactivating – RI). Chứng minh sớm nhất được thực hiện bởi Tumer và đồng nghiệp của cô, họ thí nghiệm đột biến xóa của PAP trong cây thuốc lá chuyển gen (Zoubenko et al. 1997). Mặt dầu PAPc (PAPW327stop) và PAPn (PAPG75D) không thể loại purin tại vòng S/R, chúng vẫn kháng được mầm bệnh thực vật. Gần đây, cơ chế ngăn chặn của RIP chống lại virus suy giảm miễn dịch ở người (human immunodeficiency virus - HIV) tái bản, mọi người đều tin rằng có liên quan đến hoạt động RI, cũng đã được readdress (Wang et al. 2003). Phương pháp đột biến điểm định hướng (Site-directed mutagenesis) sử dụng trichosanthin đã chứng minh được hoạt động RI cổ điển giải thích không đầy đủ hoạt đọng kháng HIV. Trichosanthin đột biến xóa C-terminal hay thêm amino acid giữ lại hầu hết tất cả hoạt động RI, nhưng không có hoạt động kháng HIV. Hơn nửa, ngày càng tăng bằng chứng chứng minh rằng RIP có khả năng nhận ra và hành động ngay lập tức trên virus và acid nucleotid cảm ứng virus, hủy bỏ những lây nhiêm của chúng (Rajamohan et al. 1999; Park et al. 2004). Chúng cũng được thông báo rằng hoạt động RI của RIP không được coi như chất độc tế bào (Park et al. 2002). Những kết quả này (Hình 1) cho biết (i) hoạt động enzyme của RIP chưa giải thích đầy đủ hành động bảo vệ chống lại mầm bệnh và (ii) cơ chế bảo vệ có thể bắt đầu bằng trực tiếp lên mầm bệnh xâm nhiễm mục tiêu hơn là ribosome vật chủ.[/FONT]
- [FONT="]Vai trò sinh học khác có thể của RIPs thu được từ bằng chứng ngày càng tăng rằng chúng có thể nhắm mục tiêu không phải ribosome. Phần còn lại adenine bảo tồn phổ biến A4324 trong vòng S/R trong một thời gian dài được coi là vị trí mục tiêu duy nhất của RIP, nhưng nghiên cứu gần đây phát hiện có thể thay thế khác vòng S/R. ví dụ, một số RIP cho thấy có khả năng giải phóng Adenine từ nhiều vị trí trong rRNA và nucleic acid poly(A), mRNA, tRNA, rRNA, và DNA (Barbieri et al. năm 1992, 1997; Park et al. 2004). Một vài RIP còn biểu lộ hoạt động chống lại poly(ADP-ribosyl)ated poly(ADP-ribose) polymerase (enzyme tự động sửa đổi; Barbieri et al. 2003). Khả năng này ảnh hưởng đến các chất khác cho phép RIP điều hòa chức năng quang trọng của tế bào[/FONT]
- [FONT="]RIP có thể điều chỉnh biểu hiện protein bằng mRNA mục tiêu thay cho ribosome. PAP cho thấy có thể ảnh hưởng trở lại trong biểu hiện chính mRNA của chúng trong nấm men chuyển gen (Parikh et al. 2002). Biểu hiện bên ngoài của PAP dẩn đến ổn định của chính mRNA của nó sau khi mức độ mRNA được điều chỉnh lên bằng cơ chế xảy ra độc lập với loại purine rRNA và dịch mã. Trong hệ thống thực vật, gene ME1 cảm ứng bằng xử lý jasmonic acid, nhưng mức độ phiên mã cao gene ME1 không có kết quả trong dịch mã thành protein in vitro (Vepachedu et al. 2003). Trong nghiên cứu in vitro cho thấy ME1 có thể loại purine chính mRNA của chúng và theo cách đó ức chế dịch mã chính nó. Bởi vậy, có thể rằng RIP có khả năng điều hòa mức độ biểu hiện của chính nó.[/FONT]
- [FONT="]Mặc dù đã được nghiên cứu tỉ mỹ từ lâu dài như là một độc chất hữu ích tiềm năng, vai trò và chức năng của RIP trong thực vật và sự phân bố tự nhiên vẫn còn khó hiểu. việc nhận dạng RIP từ cơ thể mô hình N. tabacum, chung với genomic và phương pháp công nghệ sinh học, vhawngr hạn như mutagenesis và gene silencing, có thể là một cách hiệu quả lấp lỗ hổng kiến thức. trong một vài năm tới, nó có khả năng phát triển nhanh trong lĩnh vực này. Dẫn đầu là làm sáng tỏ chức năng sinh học của RIP và chứng minh vai trò coe bản trong tế bào thực vật và tiềm năng đáng kể của chúng trong một vài ứng dụng chữa bệnh bao gồm chuẩn bị độc chất miễn dịch mục tiêu khối u và u máu ác tính. [/FONT]